译者注:本文翻译了Jay Hegde和David C. Van Essen的论文《Selectivity for Complex Shapes in Primate Visual Area V2》,V1区的研究已经相当透彻,IT区的很多研究也表明了物体的选择性,大家都很好奇从V1到IT区的整个object recognition过程中发生了什么,这些intermedia area中的神经元有哪些特性呢?本文是一个较好的尝试。原文地址:http://www.jneurosci.org/content/20/5/RC61.full.pdf
灵长类动物视觉皮层V2区的复杂形状选择性
结果
复杂轮廓和光栅的V2区细胞选择性

有两个例子,其中一个对复杂的几何刺激比简单的表现出了明显的偏好,如图1所示,使用颜色来标注响应度。图1A所示的细胞对直角具有最佳响应。这个细胞的适应范围很小,只有两个其他刺激(一个锐角和半圆)的反应效果能超过最大效果值的一半。从这个细胞对简单条形刺激的响应中无法预测到它会对直角刺激的产生最佳响应(63次/秒),因为这个细胞对直角组成部分中一个线条方向有较差的响应(17次/秒)而对另一个组成线条方向完全没有反应。这个细胞的形状选择性需要特别注意,因为几乎没有其他刺激可以引发它的响应,及时那些刺激包含最佳直角的两个主成分方向(其他直角刺激,十字和双曲光栅)。第三行的角和弧比第一行有更好的效果,这显示了对角度和/或曲率标志的极性选择性,而不是简单的同线性偏差。换句话说,这个细胞对每个se的几何图形,也对最佳图形的朝向有选择性。更重要的是,对这个细胞来说(同样适用于我们所有例子中的细胞),响应的强度在感受野内三个jitter(抖动)位置中每一处的刺激都是相当的一致。这说明刺激选择性与传统感受野内的非均匀性和固定窗口内眼球位置的波动无关(数据未显示)。图1B展示了另一个对复杂的轮廓刺激(包括弧线,特别是是各个朝向的四分之三弧和圆)有高度选择性的细胞,但只对每个相同刺激对中的较大的那个有选择性。与前一个例子不同的是,这个细胞对一系列光栅刺激都有较好的响应。最佳效果的光栅刺激包括同心环和高频螺旋,这与细胞的弯曲轮廓偏好相一致。
图1A和B的下方的响应直方图显示了在四个光栅子类和十个轮廓子类中的每一类中最佳刺激的平均响应SEM。相对于峰值响应和响应级别的范围而言错误的线条会有小的尺寸,这说明这些细胞的神经响应是相当一致的。相对于峰值响应和响应级别的范围而言较小的误差棒说明这些细胞的神经响应是相当一致的(同样见图1)。这些细胞也许传递了关于不同刺激之类之间形状差异的重要的信息,我们下面定量地来处理这个问题。前面的观察说明单独的V2细胞可以显式地对不同形状特性进行信息编码并且一个指定的细胞对简单的线条和光栅产生的反应无法预测其的最佳刺激和一个较大的刺激集合的整体响应情况。
形状选择性的众码分析
我们通过定量分析来解决下面这些关于单个V2神经元的形状选择性的问题:
(1)细胞的偏好刺激是简单还是几何上复杂的图形,这个偏好静态显著么?
(2)偏好每一个复杂图形子类的细胞们是统一的还是有偏差的?
(3)个体细胞们是否传达了形状特性的信息,而不是仅仅归因于刺激的朝向、尺寸和空间频率?
(4)个体细胞们是否传达了最佳刺激子类中的朝向、尺寸和/或空间频率的信息?
这些分析针对光栅和轮廓分别进行,其中更加引人注目的结构来自于对轮廓的分析。
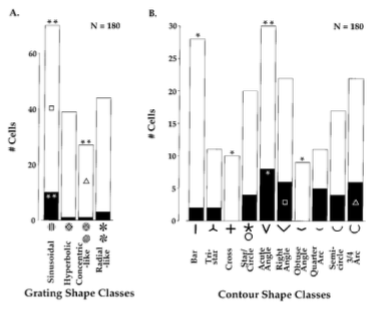
图2A中根据每个细胞的最佳光栅刺激情况指定四个光栅子类中的一个。大多数细胞(180中110个,61%)偏好非笛卡尔光栅的三个子类中的一个子类,
其中70个细胞(39%)对正弦光栅响应最好。110个中的5个对最有效的非笛卡尔光栅比最有效的简单光栅(通过Bonferoni校正后每个假设p < 0.05)响应效果要好,其发生率并不比预期的大。这非常有可能归因于许多V2细胞已经被广泛地(尽管是系统第)调制到一类光栅刺激上(见图1B)。
最有效轮廓刺激的分析如图2B所示。相比那些最有效线条刺激,偏好复杂轮廓的细胞占到较大部分(180中152个,84%)。在35个细胞中(19%),这个偏好是静态显著的。(通过Bonferoni校正后p < 0.05)。将近四分之三(152分之112,74%)的细胞相比较简单线条更加偏好复杂图形,并且相比较于最有效的正弦光栅更偏好复杂轮廓。六分之一的细胞(180中130,17%)偏好锐角。通过随机化分析确定偏好复杂轮廓的细胞整体分布和拥有显著复杂轮廓偏好的细胞的分布不是随机的(分别p<0.005和0.5)。180个中只有28个(16%)的细胞对线条刺激有最佳响应,并且只有两个细胞(1%)的响应显著好于复杂图形刺激的响应。
不同形状特性的选择性
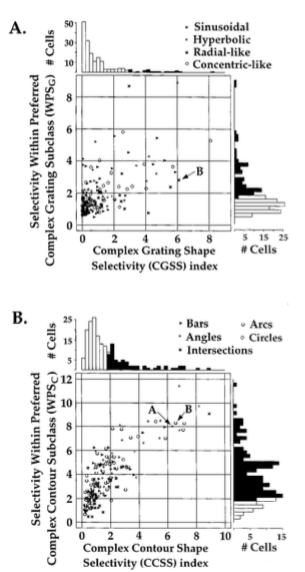
诸如图1中的细胞是否携带了形状特征的关键信息取决于对不同刺激的不同反应超过对相同刺激的不同反应的程度。如图3中,针对每个复杂刺激子类中的峰值响应(图3AB,横轴)和每个产生最大峰值响应的复杂图形子类中全部刺激(图3AB,纵轴)我们进行处理这个问题。为了确定针对不同的非笛卡尔光栅子类的指定细胞响应的调制情况,我们首先计算这个细胞对最有效的双曲、类同心和类极型光栅的响应方差,然后使用随机化分析来确定这个方差不同于随机的程度。我们计算了一个复杂光栅形状选择性指数(CGSS),定义为三个峰值响应的实际方差和平均随机方差的比率。我们也针对光栅计算了一个偏好子类内的选择性指数,定义为该细胞对子类内12个刺激的实际方差和随机方差之间的比率,包括这个细胞的最有效非笛卡尔光栅刺激。这两个指数一起用来衡量一个或多个图形特征的细胞响应调制程度。图3A展示了V2细胞对光栅刺激的调制程度。对于大约十分之一(180中19个,11%)的细胞,复杂光栅的响应方差明显高于随机方差(p<0.05),说明这些细胞能够区分非笛卡尔光栅的子类。
这19个细胞的平均CGSS值为4.7,说明响应方差是随机方差的4.7倍。对于超过三分之一的细胞(180中67个,37%),子类内响应方差明显高于随机方差(p<0.05),WPSG均值为3.1。为了分析复杂轮廓刺激的响应情况(图3B),我们计算了一个复杂轮廓细胞选择性指数(CCSS),定义为细胞对来自九个复杂轮廓刺激子类中的每个偏好刺激的响应的实际方差与平均随机方差的比率。
我们也计算了子类内的轮廓选择性指数(WPSC),定义为细胞对子类内的八个刺激的响应方差与随机方差的比率。大约三分之一的细胞(180中的62个 34%),复杂轮廓响应方差显著高于随机方差(p<0.05)。这63个细胞的CCSS平均值为3.5,意味着响应方差是随机方差的3.5倍。V2细胞也显示了子类内较强的响应调制。这些细胞中大约有五分之四(180个钟147,82%)的响应方差显著高于随机方差,平均WPSC值为4.4.对于轮廓响应,跨刺激子类选择性指数和偏好子类内的选择性指数是彼此强相关的(r=0.81;p<0.01)。52个指数超过2.0的细胞中,具有角和弧线最佳偏好的细胞是最普遍的(分别是19和15个),而偏好十字和线条的是最少的(分别是10个和8个)。
上面的结果说明许多V2细胞的响应针对复杂图形的特性并且针对朝向、尺寸和空间频率都有较大的调制。综合峰值响应分析的结果,这些结果表明许多V2细胞都充分地携带了复杂形状的详细的潜在有用的信息。
讨论
V2感受野特性的起源
V2区细胞针对复杂轮廓和光栅的选择性引发了一个非常的有趣的问题:这个感受野特性是如何产生的呢?一种可能是这些特性通过来自V1区细胞(对低维形状信息,如朝向、空间频率有选择性,见Riesenhuber and Poggio, 1999)恰当模式的上升输入产生于V2区的。
例如直角的选择性可能源于V1细胞的两个子集,每个子集只对两个相互垂直方向中的一个有选择性,这有点像当年Hubel和wiesel(1965)年指出的猫脑19区的高维复杂超级细胞。V2区复杂形状选择性也可能来自于
旁边的对简单刺激有选择性的V2区细胞和/或诸如V4的较高视觉区域的下降输入(Gallant等, 1996)。最后,V2细胞的特征也许反应了V1细胞自身的复杂细胞偏好。这可以产生于nonclassical surround interactions(非经典环绕的交互作用)(Knierim and Van Essen, 1992; Lamme, 1995; Das and Gilbert, 1999)或者
来自于经典感受野内部的复杂细胞选择性(Versavel等, 1990; Hegd ́e and Van Essen, 1999)。